Glukagon er et peptidhormon, produsert av alfa-celler i bukspyttkjertelen. Glukagon øker konsentrasjonen av glukose og fettsyrer i blodet, og regnes som det viktigste katabolske hormonet i kroppen. Glucagon brukes også som medisin for å behandle en rekke helsemessige forhold. Effekten av glukagon er motsatt den av insulin, som senker ekstracellulær glukose. Glucagon er produsert fra proglucagon, kodet av GCG-genet.
Bukspyttkjertelen frigjør glukagon når mengden glukose i blodet er for lav. Glukagon får leveren til å delta i glykogenolyse: omdanne lagret glykogen til glukose, som frigjøres i blodet. Høye blodsukkernivåer stimulerer derimot frigjøringen av insulin. Insulin gjør at glukose kan tas opp og brukes av insulinavhengig vev. Dermed er glukagon og insulin en del av et tilbakemeldingssystem som holder blodsukkernivået stabilt. Glucagon øker energiforbruket og er forhøyet under stressforhold. Glucagon tilhører secretin-familien av hormoner.
Funksjon av glukagon
Glukagon hever generelt konsentrasjonen av glukose i blodet ved å fremme glukoneogenese og glykogenolyse. Glukagon reduserer også fettsyresyntese i fettvev og lever. Glucagon fremmer lipolyse i disse vevene, noe som får dem til å frigjøre fettsyrer i sirkulasjon der de kan kataboliseres for å generere energi i vev som skjelettmuskulatur når det er nødvendig.
Glukose lagres i leveren i form av polysakkaridglykogenet, som er en glukan (en polymer som består av glukosemolekyler). Leverceller (hepatocytter) har glukagonreseptorer. Når glukagon binder seg til glukagonreseptorene, omdanner levercellene glykogen til individuelle glukosemolekyler og frigjør dem i blodet, i en prosess kjent som glykogenolyse. Når glukoselagring blir utarmet, oppmuntrer glukagon leveren og nyrene til å syntetisere ekstra glukose ved glukoneogenese. Glukagon slår av glykolyse i leveren, noe som fører til at glykolytiske mellomprodukter skyves til glukoneogenese.
Glucagon regulerer også hastigheten på glukoseproduksjon gjennom lipolyse. Glucagon induserer lipolyse hos mennesker under insulinundertrykkelse (for eksempel diabetes mellitus type 1).
Glukagonproduksjon ser ut til å være avhengig av sentralnervesystemet gjennom veier som ennå ikke er definert. Hos virvelløse dyr er det rapportert at fjerning av øyestikk påvirker glukagonproduksjonen. Å fjerne øyestjelen hos unge kreps produserer glukagonindusert hyperglykemi.
Virkningsmekanisme for glukagon

Glukagon binder seg til glukagonreseptoren, en G-proteinkoblet reseptor, som ligger i plasmamembranen i cellen. Konformasjonsendringen i reseptoren aktiverer G-proteiner, et heterotrimert protein med α-, β- og γ-underenheter. Når G-proteinet samhandler med reseptoren, gjennomgår det en konformasjonsendring som resulterer i erstatning av BNP-molekylet som var bundet til α-underenheten med et GTP-molekyl. Denne substitusjonen resulterer i frigjøring av α-underenheten fra β- og γ-underenhetene. Alfa-underenheten aktiverer spesifikt det neste enzymet i kaskaden, adenylatsyklase.
Adenylatsyklase produserer syklisk adenosinmonofosfat (syklisk AMP eller cAMP), som aktiverer proteinkinase A (cAMP-avhengig proteinkinase). Dette enzymet aktiverer i sin tur fosforylase-kinase, som deretter fosforylerer glykogenfosforylase b (PYG b), og omdanner det til den aktive formen kalt fosforylase a (PYG a). Fosforylase a er enzymet som er ansvarlig for frigjøring av glukose 1-fosfat fra glykogenpolymerer.
Et eksempel på veien ville være når glukagon binder seg til et transmembranprotein. De transmembrane proteiner samhandler med Gɑβ𝛾. G2 skiller seg fra Gβ2 og samhandler med transmembranproteinet adenylylsyklase. Adenylylsyklase katalyserer omdannelsen av ATP til cAMP. cAMP binder seg til proteinkinase A, og den komplekse fosforylerer fosforylase-kinase. Fosforylerte fosforylase kinase fosforylerer fosforylase. Fosforylerte fosforylaser klipper glukosenheter fra glykogen som glukose 1-fosfat.
I tillegg justeres den koordinerte kontrollen av glykolyse og glukoneogenese i leveren av fosforyleringstilstanden til enzymene som katalyserer dannelsen av en kraftig aktivator av glykolyse, kalt fruktose 2,6-bisfosfat. Enzymproteinkinase A (PKA) som ble stimulert av kaskaden initiert av glukagon, vil også fosforylere en enkelt serinrest av den bifunksjonelle polypeptidkjeden som inneholder både enzymene fruktose 2,6-bisfosfatase og fosfofruktokinase-2. Denne kovalente fosforylering initiert av glukagon aktiverer førstnevnte og hemmer sistnevnte. Dette regulerer reaksjonskatalyserende fruktose 2,6-bisfosfat (en kraftig aktivator av fosfofruktokinase-1, enzymet som er det primære regulatoriske trinnet for glykolyse) ved å bremse hastigheten for dets dannelse, og derved hemme strømmen av glykolyseveien og tillate glukoneogenese å dominere. Denne prosessen er reversibel i fravær av glukagon (og dermed tilstedeværelsen av insulin).
Glukagonstimulering av PKA inaktiverer også det glykolytiske enzymet pyruvatkinase i hepatocytter.
Fysiologi
Glukagonproduksjon
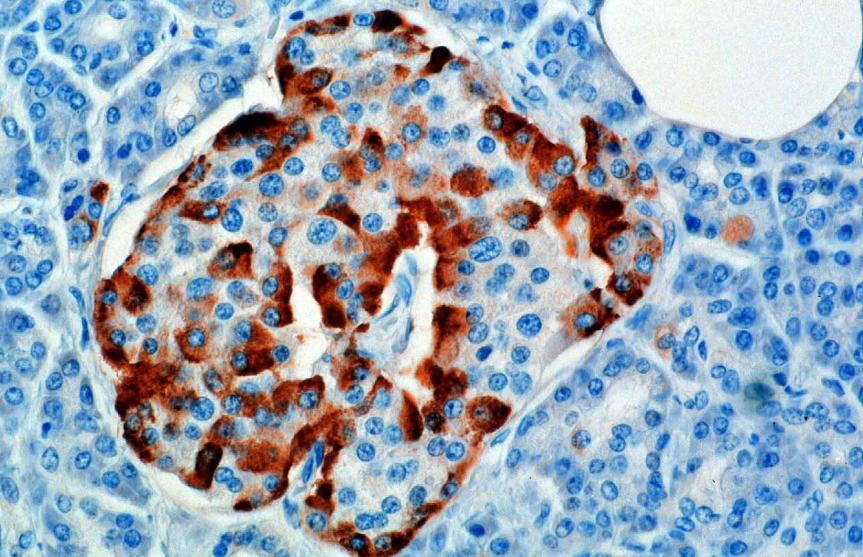
Hormonet syntetiseres og skilles ut fra alfa-celler (α-celler) i øyene i Langerhans, som ligger i den endokrine delen av bukspyttkjertelen. Produksjon, som ellers er friløpende, undertrykkes / reguleres av amylin, et peptidhormon co-utskilt med insulin fra bukspyttkjertelen β-celler. Etter hvert som plasmaglukosenivået trekker seg tilbake, reduserer den påfølgende reduksjonen i amylinsekresjon undertrykkelsen av α-cellene, slik at glukagonsekresjon muliggjøres.
Hos gnagere er alfacellene plassert i den ytre kanten av holmen. Menneskelig holmstruktur er mye mindre segregerte, og alfaceller fordeles gjennom holmen i nærheten av betaceller. Glukagon produseres også av alfaceller i magen.
Nyere forskning har vist at glukagonproduksjon også kan finne sted utenfor bukspyttkjertelen, med tarmen som det mest sannsynlige stedet for glukagon-syntese utenom bukspyttkjertelen.
Glukagon regulering
Sekresjon av glukagon stimuleres av:
- Hypoglykemi
- Epinefrin (via β2-, α2- og α1-adrenerge reseptorer)
- Arginin
- Alanin (ofte fra muskelavledet pyruvat / glutamattransaminering
- Acetylkolin
- Kolecystokinin
- Gastrisk hemmende polypeptid
Sekresjon av glukagon hemmes av:
- Somatostatin
- Amylin
- Insulin (via GABA)
- PPARy / retinoid X reseptor heterodimer.
- Økt frie fettsyrer og ketosyrer i blodet.
- Økt ureaproduksjon
- Glukagon-lignende peptid-1
Glukagon struktur
Glucagon er et 29-aminosyre-polypeptid. Dens primære struktur hos mennesker er: NH2-His-Ser-Gln-Gly-Thr-Phe-Thr-Ser-Asp-Tyr-Ser-Lys-Tyr-Leu-Asp-Ser-Arg-Arg-Ala-Gln-Asp-Phe-Val-Gln-Trp -Leu-Met-Asn-Thr-COOH.
Polypeptidet har en molekylvekt på 3485 dalton. Glukagon er et peptid (ikke-steroid) hormon.
Glukagon genereres fra spaltingen av proglukagon ved proproteinkonvertase 2 i α-celler i bukspyttkjertelen. I intestinale L-celler spaltes proglukagon til de alternative produktene glicentin, GLP-1 (et inkretin), IP-2 og GLP-2 (fremmer tarmvekst).
Patologi
Unormalt forhøyede nivåer av glukagon kan være forårsaket av bukspyttkjerteltumorer, slik som glukagonoma, hvis symptomer inkluderer nekrolytisk vandrende erytem, reduserte aminosyrer og hyperglykemi. Det kan forekomme alene eller i sammenheng med multipel endokrin neoplasi type 1.
Forhøyet glukagon er den viktigste bidragsyteren til hyperglykemisk ketoacidose ved udiagnostisert eller dårlig behandlet type 1-diabetes. Etter hvert som betacellene slutter å fungere, er insulin og bukspyttkjertel-GABA ikke lenger tilstede for å undertrykke det friløpende resultatet av glukagon. Som et resultat frigjøres glukagon maksimalt fra alfacellene, noe som forårsaker en rask nedbrytning av glykogen til glukose og rask ketogenese. Det ble funnet at en undergruppe av voksne med type 1-diabetes i gjennomsnitt tok 4 ganger lengre tid å nærme seg ketoacidose når den ble gitt somatostatin (hemmer glukagonproduksjonen) uten insulin. Å hemme glukagon har vært en populær idé om diabetesbehandling, men noen har advart om at dette vil gi sprø diabetes hos pasienter med tilstrekkelig stabil blodsukker.
Fraværet av alfaceller (og dermed glukagon) antas å være en av hovedpåvirkningene i den ekstreme flyktigheten av blodsukker i omgivelsene til en total pankreatektomi.
Historie
På 1920-tallet studerte Kimball og Murlin bukspyttkjertelekstrakter, og fant et ekstra stoff med hyperglykemiske egenskaper. De beskrev glukagon i 1923. Aminosyresekvensen til glukagon ble beskrevet på slutten av 1950-tallet. En mer fullstendig forståelse av dens rolle i fysiologi og sykdom ble ikke etablert før på 1970-tallet, da en spesifikk radioimmunoanalyse ble utviklet.
.
Discussion about this post